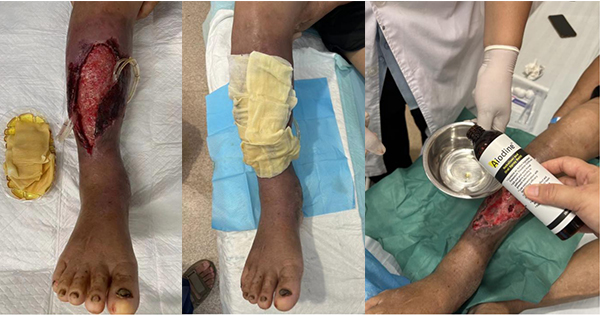
Wound healing is a dynamic and intricate biological process initiated by the body to restore damaged tissue (Gonzalez et al, 2016), regulated by various cellular and molecular mediators, including platelets, neutrophils, keratinocytes, and interleukins (Werner and Grose, 2003). The interaction between these various cell types and cytokines ensures proper progression along the wound-healing trajectory (Mahmoud et al, 2024).
Additionally, wound healing is influenced by intrinsic factors such as perfusion and oxygenation, and extrinsic factors, including nutrition, infection, and certain medications, all of which affect cellular function and repair efficiency (Beyene et al, 2020). Consequently, disruptions in signalling pathways resulting from intrinsic or extrinsic influences such as medications, including steroids, can impair intercellular communication and impede the healing process (Beyene et al, 2020; Mahmoud et al, 2024).
Steroids are potent immunosuppressive and anti-inflammatory medications that are widely used in treating various clinical conditions across different settings (Perretti and Ahluwalia, 2000). Recent evidence indicates that steroids not only alleviate symptoms of inflammatory diseases but also offer long-term therapeutic benefits, enhancing patient prognosis (Perretti and Ahluwalia 2000). Although the therapeutic efficacy of corticosteroids is well-established for certain conditions, clinical reports and animal studies have indicated that high doses may impair wound healing (Wang et al, 2013).
This article will specifically examine the effects of corticosteroids across phases of wound healing.
Material and methods
A structured literature search review was carried out to identify relevant evidence on the effects of corticosteroids on wound healing. The primary database used was Google Scholar due to its broad coverage of biomedical and clinical literature.
The keywords applied were corticosteroid, steroid, and wound healing. These terms were used in combination with Boolean operators. To maximise retrieval of relevant sources, synonyms were linked with ‘OR’ (e.g. “corticosteroids OR steroid”) and concepts were linked with ‘AND’, e.g. (corticosteroids OR steroid) AND wound healing).
Articles were included if they:
- Reported primary data or systematic evidence on corticosteroid administration and its impact on wound healing.
- Provided clinically relevant supporting information such as reviews or guidelines.
Studies were excluded if they:
- Focused solely on unrelated steroid uses (e.g. asthma, rheumatology) without addressing wound outcomes.
- Were not available in English.
The search retrieved 35 relevant articles. All titles, abstracts, and full-text articles were screened for inclusion by a single researcher (the author). Data extraction was also performed independently by the same researcher. Information was extracted on study design, population, type and dose of corticosteroid, wound type, and reported outcomes. Evidence from primary studies was synthesised with findings from secondary literature to evaluate both potential beneficial and harmful effects of corticosteroid use in wound healing.
A narrative synthesis was used rather than meta-analysis due to heterogeneity in study designs and outcome measures. As both screening and data extraction were conducted by a single reviewer, this represents a methodological limitation that may introduce selection or interpretation bias. However, this limitation was partly addressed by applying clear inclusion criteria and selecting studies that directly supported the review focus.
Results
The structured search yielded 35 relevant articles, of which 19 were primary research studies and 16 were reviews or supporting literature. Findings were grouped according to their impact on different stages of wound healing.
Inflammatory phase
Laboratory and preclinical studies have shown that corticosteroids delay the initiation of the inflammatory phase by downregulating cytokine and chemokine expression, thereby reducing the recruitment of inflammatory cells (Hübner et al, 1996; Bhattacharyya et al, 2007; Chatzopoulou et al, 2016; Xie et al, 2019). They also inhibit macrophage differentiation into the pro-inflammatory M1 phenotype (Xie et al, 2019; Kim et al, 2020; Öztürk 2023). This is a key process in the early inflammatory response, which contributes to a delayed transition to the proliferative phase.
Proliferative phase
Corticosteroid exposure was consistently associated with reduced angiogenesis and fibroblast proliferation leading to delay in functional granulation tissue formation (Hein et al, 1988; Nauck et al, 1998; Nguyen et al, 2022; Anker et al, 2023).
Remodelling phase
Multiple studies described adverse effects of corticosteroids on collagen deposition and cross-linking with evidence indicated that corticosteroid-treated wounds had lower tensile strength compared to those untreated with corticosteroids, reflecting compromised collagen maturation, thereby increasing the risk of wound dehiscence (Dostal and Gamelli, 1990; Oishi et al, 2002; AlbertiI et al, 2012).
Discussion
Inflammatory phase
Following tissue injury, various inflammatory cytokines, such as interleukin 1-alpha (IL-1α) and -beta (IL-1β) and tumour necrosis factor alpha (TNF-α), are released to modulate inflammation by promoting immune cell recruitment, tissue proliferation, re-epithelialisation and tissue remodelling (Mahmoud et al, 2024). One way steroids affect wound healing is by suppressing the expression of these pro-inflammatory cytokines, which reduces the signalling molecule needed to recruit other inflammatory cells and mediators to the site of injury (Ehrchen et al, 2019).
Hübner et al (1996) conducted an in vivo study in mice to examine the effects of glucocorticoids on the regulation of pro-inflammatory cytokines during wound healing and found a significant increase in IL-1α, IL-1β and TNF-α expression following tissue injury. However, this elevation was markedly reduced in glucocorticoid-treated mice, suggesting that steroid treatment effectively suppresses the expression of these cytokines.
Similarly, Bhattacharyya et al (2007) reported findings consistent with Hübner et al (1996) in an in vitro study. They showed that glucocorticoids selectively inhibit p38 MAPK activation, leading to a reduction in pro-inflammatory cytokine production, particularly TNF-α and IL-6. In this study, dexamethasone treatment led to a substantial reduction in cytokine secretion in normal macrophages, with TNF-α and IL-6 levels decreased by 80% and 90%, respectively. In contrast, glucocorticoid receptor macrophage knock-out mice showed no significant change in cytokine levels following treatment, indicating a loss of glucocorticoid responsiveness due to the absence of functional receptors.
Both the in vitro and in vivo studies demonstrated that steroid treatment potentially disrupts wound healing by reducing cytokine expression following inflammatory stimuli or tissue injury (Hübner et al, 1996; Bhattacharyya et al, 2007). The in vitro findings highlighted a direct immunosuppressive effect, while the in vivo results confirmed systemic modulation in a physiologically relevant animal model. Despite these shared outcomes, the studies differed in methodology, particularly in steroid dosage and timing. The in vitro study involved a single steroid dose administered 3 hours before lipopolysaccharide exposure, whereas the in vivo study employed a prolonged regimen, with daily steroid administration 3 days prior to injury and continued for 5 days post-wounding. This variation in dosing and treatment duration may influence the observed effects and complicate clinical translation.
Furthermore, differences in steroid protocols, potential in vivo confounding factors and the lack of human data emphasise the need for further research to ensure the clinical relevance of these findings (Schulze et al, 1997). Although current findings suggest that steroids may disrupt wound healing by suppressing pro-inflammatory cytokine expression, critical for immune cell recruitment, these effects must be confirmed through rigorous human studies employing standardised dosing and treatment protocols to ensure clinical relevance.
Other studies have demonstrated that steroids impair the recruitment of inflammatory cells, particularly neutrophils and macrophages, to the injury site by downregulating chemokines, such as interleukin-8 (IL-8) and CCL2, which serve as key chemoattractants (Xie et al, 2019). Consequently, the reduced presence of these immune cells may compromise the initiation and maintenance of the inflammatory phase. While this promotes a more controlled resolution of inflammation, the reduced inflammatory response may disrupt the wound healing process by suppressing the early inflammatory phase, potentially hindering effective pathogen clearance (Landén et al, 2016).
This is supported by a study conducted by Chatzopoulou et al (2016), who investigated the immunomodulatory effects of glucocorticoids on inflammatory cell migration in zebrafish, focusing on leukotrienes as the key chemoattractant for neutrophil recruitment. Using ELISA, the study demonstrated that tail fin amputation in zebrafish led to an approximately threefold increase in leukotriene levels. However, this increase in leukotriene levels was suppressed by beclomethasone, likely by affecting post-transcriptional processes or enzymatic activity involved in leukotriene biosynthesis, thereby reducing neutrophil migration.
Interestingly, the researchers also found that while neutrophils and macrophages migrate to the wound site, they exhibited distinct responses to glucocorticoids. Beclomethasone specifically reduced neutrophil migration without affecting macrophage recruitment. Quantification of neutrophils in the tail fin revealed no significant difference in total neutrophil count between treated and untreated larvae, suggesting that beclomethasone inhibits neutrophil migration rather than reducing neutrophil numbers.
However, while zebrafish embryos offer the advantage of direct visualisation of immune cell migration due to their optical transparency and share a highly conserved innate immune system similar to humans, their poikilothermic nature, which affects metabolic activity and cytokine signalling, may alter the pharmacological mechanisms of certain medication, including temperature-sensitive steroids (Dudley et al, 2012; Grada et al, 2018; Naomi et al, 2021). Therefore, caution is required when translating data from zebrafish in vivo studies to human clinical practice, necessitating further validation in mammalian models.
A study by Xie et al (2019) confirmed Chatzopolou et al’s findings. Using qPCR analysis, they revealed that beclomethasone suppressed the expression of IL-8 and Cxcl-18b, which are essential for neutrophil recruitment. In contrast, beclomethasone had no effect on Ccl2 and Cxcl-11a, which regulate macrophage migration. These findings suggest that glucocorticoids selectively inhibit neutrophil recruitment by downregulating neutrophil-specific chemoattractant genes (Zentay et al, 1999), while leaving macrophage migration unaffected (Xie et al, 2019). Clinically, this targeted modulation potentially helps control excessive neutrophilic inflammation in chronic non-healing wounds while preserving macrophage-mediated tissue repair and homeostasis (Gauthier et al, 2018).
Despite previous findings emphasising the inflammatory role of macrophages in wound healing, Xie et al (2019) demonstrated that glucocorticoids impair this process by suppressing macrophage differentiation into the pro-inflammatory M1 phenotype, which is crucial during the early inflammatory phase while largely sparing M2 marker expression. RNA-sequencing analysis revealed downregulation of M1-associated genes, including IL-6, IL-1β, and TNF-α, compared to the amputation-only group, suggesting a selective inhibition of M1 polarisation (Xie et al, 2019).
This finding is consistent with Kim et al (2020), who also reported that glucocorticoids inhibit macrophage polarisation away from M1. In parallel, Ozturk (2023) observed a shift towards the anti-inflammatory M2 phenotype, which may dampen necessary early inflammation. While such modulation may benefit pathological contexts involving excessive or chronic inflammation, it risks dampening the initial immune activation required for effective pathogen clearance and debris removal in normal wound healing.
Proliferative phase
The proliferative phase marks the stage where angiogenesis, granulation tissue formation and wound re-epithelialisation occur (Singh et al, 2017). In 1998, Nauck et al. investigated the anti-angiogenic effects of glucocorticoids using cultured human aortic vascular smooth muscle cells. Their findings demonstrated that corticosteroids suppressed platelet derived growth factor (PDGF)-induced vascular endothelial growth factor (VEGF) gene expression in a dose-dependent manner, indicating a potential mechanism by which glucocorticoids inhibit angiogenesis (Nauck et al, 1998).
Nguyen et al (2022) conducted an in vivo study to investigate the impact of a mineralocorticoid receptor (MR) antagonist on wound angiogenesis in steroid-pretreated mice. At day 5 post-wounding, blood vessel density, measured by CD31+ staining, was significantly reduced in the steroid-treated group. However, administration of the MR antagonist restored CD31+ expression by 77% (p<0.05) and reversed the suppression of key pro-angiogenic factors, including VEGF-A, FGF2, and CXCL12. These findings suggest that steroid-induced MR activation impairs angiogenesis, while MR antagonism effectively mitigates this effect.
However, although blood vessel density is commonly used to assess angiogenesis, as demonstrated in this study, it primarily reflects the quantity of newly formed vessels rather than their functional maturity or perfusion capacity (Hoeben et al, 2004). Many nascent vessels formed during the early stages of wound healing are non-perfused and lack functionality (Hoeben et al, 2004). Therefore, relying solely on vessel density may not accurately reflect the overall effectiveness of angiogenesis.
While in vitro studies have shown that steroids can inhibit blood vessel formation by suppressing VEGF gene expression (Nauck et al, 1998), their effect on vessel functionality in clinical settings should be evaluated using parameters capable of measuring vessel perfusion, such as laser Doppler perfusion imaging (Aarnink et al, 2009). This is crucial, as perfusion directly reflects the vessel’s ability to deliver metabolic nutrients and remove waste products from the surrounding tissue (Aarnink et al, 2009).
Other studies have shown that corticosteroids influence fibroblast activity during the proliferative phase of wound healing (Beer et al, 2000). Hein et al (1988) investigated the effects of three corticosteroids (desoximetasone, hydrocortisone, and prednicarbate) on human dermal fibroblasts. In their study, cultured fibroblasts derived from human skin biopsies were exposed to varying concentrations of each corticosteroid. The results revealed that desoximetasone and hydrocortisone significantly inhibited fibroblast proliferation by up to 50% compared to untreated controls without affecting cell morphology or viability. Conversely, prednicarbate did not exhibit a comparable inhibitory effect. Additionally, even at low concentrations, potent corticosteroids reduced fibroblast chemotaxis, suggesting that the impact of corticosteroids on fibroblast function is both dose-dependent and derivative-specific.
While the absence of detailed patient characteristics in the methodology may enhance the generalisability of the study by not restricting the findings to a specific patient subgroup (Filbey et al, 2023), it simultaneously limits the interpretability of the results, as variables such as age or comorbidities are known to affect skin cell behaviour independently and could act as confounding factors in evaluating the actual effect of steroid treatment (Beyene et al, 2020).
Although a general impairment of fibroblast migration is often attributed to corticosteroid exposure, this appears to contrast with findings by Anker et al (2023), who reported no statistically significant differences in migration between fibroblasts incubated with triamcinolone-treated seroma and those without (p<0.364). In their in vitro model, human dermal fibroblasts isolated from pannus tissue were cultured in seroma-conditioned media, either with or without triamcinolone. Interestingly, they observed that fibroblasts exposed to early postoperative seroma (<10 days) exhibited slower migration when residual steroid levels were presumably higher. While not statistically significant overall, this observation suggests a concentration-dependent, time-sensitive effect.
Such variability may be attributable to pharmacokinetic differences in steroids, which exhibit dose- and time-dependent clearance dynamics influenced by factors like plasma protein binding (Rohatagi et al, 1997). McMaster et al (2008) emphasised that single time-point assessments may fail to capture the complexity of steroid responses, advocating for real-time analytical approaches to elucidate dynamic cellular behaviour.
Remodelling phase
The remodelling phase of wound healing is characterised primarily by the rearrangement and cross-linking of collagen fibres, which restore tissue tensile strength and long-term structural integrity (Mahmoud et al, 2024). One widely accepted metric for assessing the success of wound remodelling is tensile strength, which reflects the mechanical resilience of the newly formed tissue (Gonzalez et al, 2016).
Dostal and Gamelli (1990) conducted a study using a mechanical testing method to measure wound disruption strength in mice. The study compared the effects of three types of corticosteroids (methylprednisolone, dexamethasone, and hydrocortisone) and observed a dose-dependent reduction in tensile strength across all groups. Notably, dexamethasone and hydrocortisone significantly weakened the wounds at all tested doses (p<0.05), while methylprednisolone only led to a significant decrease at higher doses. These findings suggest that although all corticosteroids may negatively impact wound remodelling, their relative toxicity varies.
To investigate the molecular basis of altered collagen metabolism during the remodelling phase, Oishi et al (2002) used a rat skin model and found that dexamethasone markedly suppressed mRNA expression of type I and III collagen, significantly reducing tropocollagen synthesis (p<0.01), a precursor of mature collagen fibres. Type III collagen appeared more sensitive to suppression than type I. Additionally, dexamethasone downregulated key enzymes involved in collagen turnover, including collagenase and tissue inhibitors of metalloproteinases (TIMPs), disrupting extracellular matrix remodelling (Oishi et al, 2002). This dual suppression of synthesis and degradation likely impairs matrix integrity and contributes to reduced wound tensile strength. However, the 1 mg/kg subcutaneous dose used, equivalent to approximately 11 mg/day in human based on body surface area (BSA) conversion (Nair and Jacob 2016), represents a high-end clinical exposure. While suitable for investigating mechanistic effects, extrapolation to routine therapeutic contexts warrants caution due to interspecies pharmacodynamic differences.
A similar finding was reported by Albertil et al (2012), who demonstrated that mice treated with local and systemic hydrocortisone exhibited a significant reduction in scar resistance on postoperative day 7 (p<0.005). However, no significant differences were observed on days 14 and 21 (p>0.005). This transient impairment suggests that corticosteroids primarily delay tissue remodelling rather than permanently inhibit it.
The early-phase weakness likely reflects suppressed fibroblast activity, diminished collagen synthesis, and impaired angiogenesis (Oishi et al, 2002) induced by corticosteroids. However, the absence of significant differences at later time points (days 14 and 21) indicates that the healing process may eventually recover (AlbertiI et al, 2012), implying that corticosteroid effects are temporary and potentially reversible. Importantly, while tissue strength may recover over time, this temporal vulnerability could increase clinical risks such as wound dehiscence or infection in the early postoperative period, particularly in surgical settings.
Wang et al (2013) highlighted the risk of compromised tissue integrity, reporting an approximate 30% reduction in wound tensile strength in animal models treated with high-dose corticosteroids (15–40 mg/kg/day). In contrast, a human randomised controlled trial by Schulze et al (1997) found no significant difference in wound healing between corticosteroid-treated and control groups following acute high-dose administration. This apparent discrepancy highlights the need to consider species differences, dosing duration, and pharmacodynamic variability when evaluating corticosteroid effects on wound remodelling.
Although corticosteroids may initially weaken tissue by suppressing collagen synthesis and matrix remodelling, these effects appear reversible over time (AlbertiI et al, 2012). With careful timing and dosage, corticosteroid therapy may remain clinically acceptable (Schulze et al, 1997), although caution is warranted during periods of increased mechanical stress or infection risk.
Conclusion
Corticosteroids remain valuable therapeutic agents because of their potent anti-inflammatory effects, yet their potential to impair wound healing across all phases raises important concerns for clinical practice. Their impact appears to vary depending on drug type, dose, and duration of administration. Further human-based studies are needed to clarify these effects and to establish safe prescribing practices that minimise harm to wound outcomes
Given the suppressive effects of corticosteroids across all phases of wound healing, clinicians should exercise particular caution when prescribing them to patients with acute wounds. Early corticosteroid exposure can disrupt cytokine signalling and impair immune cell recruitment during the inflammatory phase. Therefore, delaying corticosteroid initiation until this phase has progressed may help mitigate the risk of impaired healing. The observed reduction in collagen synthesis and tensile strength, particularly during the remodelling phase, underscores the importance of close monitoring for wound dehiscence, infection, and delayed healing in patients receiving corticosteroids. Importantly, not all corticosteroids exert uniform effects, as evidence suggests their toxicity varies according to the specific agent, dosage, and treatment duration.
As most current findings are derived from animal models or in vitro studies, further human research using standardised dosing protocols and validated outcome measures, is essential to guide clinical practice. In the interim, clinicians should interpret preclinical evidence with caution and individualise treatment decisions based on patient-specific factors.